


Ce dossier écrit est la retranscription écrite de l’épisode 372 de Podcast Science, co-écrit par Pierre Kerner (1ère partie) et Eléa Héberlé (2ème partie)
Pierre Kerner vous explique le sexe :
Il y a peu de temps, naissait Isaac Alexander Smythe Kerner, mon neveu, le fils de Jacques qui était venu vous parler de la physique des Jeux Vidéos pour l’épisode 369. Et jusqu’au dernier moment je ne savais pas, du souhait de mon frère, si j’allais de nouveau avoir une nièce ou pas. Mais ça y’est, la génétique a parlé : Isaac est porteur d’un grand chromosome X et d’un petit chromosome Y dans le noyau de ses cellules. Et c’est ce petit chromosome Y qui a déterminé le devenir d’une partie de ses cellules embryonnaires qui ont permis de le fournir en deux belles gonades qui pendent à l’extérieur de son corps. Je profite donc de cette introduction pour souhaiter longue vie à mon neveu Isaac et le remercier de ce prétexte parfait pour introduire un sujet passionnant : celui de la détermination sexuelle.
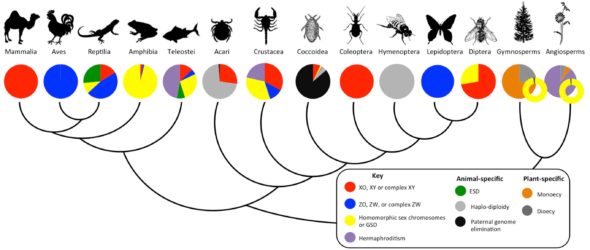
Hormis exceptions rarissimes et sans entrer dans les questions de genres, on peut facilement affirmer que c’est la présence ou non du chromosome Y qui détermine le sexe d’un enfant, dans le sens où il va contrôler le développement de testicules pendant l’embryogenèse. Sachant que d’un point de vue évolutif, la seule population de cellules humaines qui participe à la génération ultérieure est celle qui permet la formation d’ovules ou de spermatozoïdes, on peut comprendre à quel point le développement des organes permettant la production et le maintien de ces cellules est crucial pour le succès évolutif de l’espèce. Du coup il est assez surprenant de découvrir, pour peu qu’on s’y intéresse, l’incroyable diversité de détermination sexuelle qui existe chez les êtres vivants
Comme nous vous l’avions dévoilé lors de l’émission 299, il existe des gènes qui permettent de contrôler le développement d’un œil et qui sont conservés entre des espèces très distinctes comme par exemple une mouche et une souris. Par contre, il ne semble pas exister de réseaux de gènes de la détermination sexuelle qui soit universel.
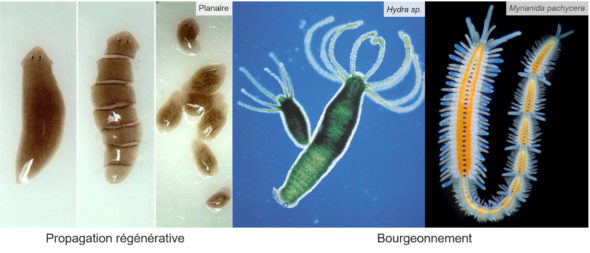
Une piste d’explication est à trouver déjà dans les types de reproductions dont sont capables les organismes vivants. Chez les animaux par exemple, le fait qu’une espèce soit caractérisée par deux sexes distincts, ce qu’on appelle le gonochorisme, c’est loin d’être la règle. Certaines espèces n’ont pas de sexes distincts et se reproduisent par divers processus asexués (bourgeonnement, division, parthénogenèse, etc.)…
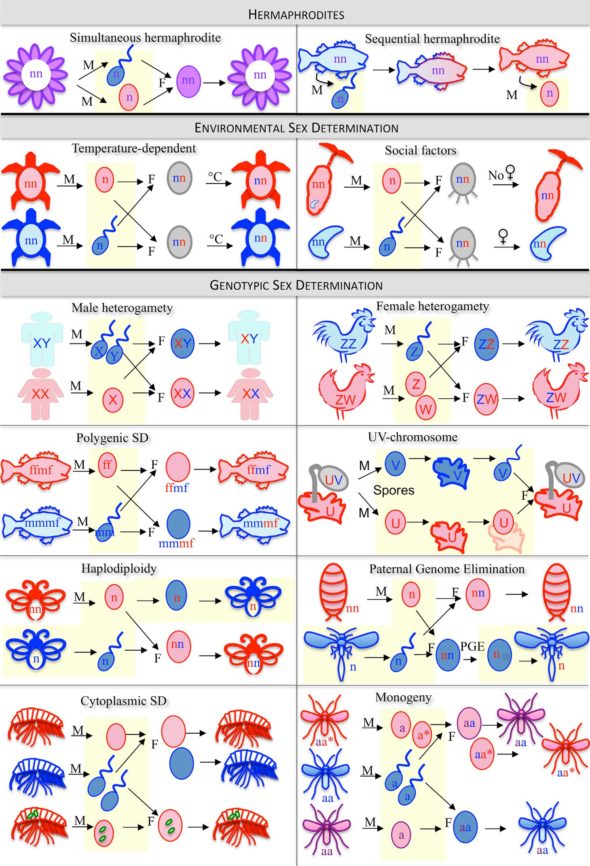
…alors que d’autres ne s’embarrassent pas de choisir et possèdent deux sexes simultanément (ce qu’on appelle l’hermaphrodisme).
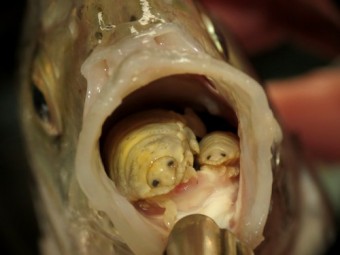
Dans mon livre “Moi, Parasite”, je présente même le cas d’une espèce parasitaire de crustacé, Cymothoa exigua, qui pratique l’hermaphrodisme séquentiel, car les individus commencent leur vie en étant tous mâles, puis selon l’occupation de leur hôte, se transformeront en femelles si celui-ci est inoccupé, ou resteront mâles si une femelle si trouve déjà. Et si la femelle vient à mourir, c’est le signal pour le mâle qu’il faut changer de sexe. Tout ça sachant que l’hôte est un poisson dont la cavité buccale se transforme en chambre nuptiale, ces parasites étant adeptes de leur langue qu’ils remplacent progressivement…
Mais chez leurs hôtes, comme le poisson clown, ce genre d’hermaphrodisme séquentiel existe aussi. Les poissons clowns ce sont ceux qui ont inspiré le film d’animation “Le Monde de Némo”. Beaucoup de personnes ont appris l’incroyable vérité sur le papa de Némo en découvrant la BD de Marion Montaigne sur son blog “Tu mourras moins bête”.


Attention Spoilers : à la mort de la maman de Némo, Marin, le papa aurait dû… devenir la nouvelle maman de Némo. En effet, chez les poissons clown, il y a une structure sociale très stricte : une seule femelle par groupe (le plus gros poisson), un mâle reproducteur et des petits juvéniles interdits de reproduction. Si la femelle vient à mourir, le mâle change de sexe et devient la nouvelle femelle et un des juvéniles peut prendre le rôle de mâle. Cela signifie donc que tous les poissons clowns naissent mâles, mais que seules les circonstances sociales peuvent décider d’un changement de sexe pour devenir femelle.
Un autre type de déterminisme sexuel qui me fascine, c’est celui qui est uniquement tributaire de l’environnement. Ce déterminisme est définitif, il n’y a pas de changement de sexe possible, mais contre il est déterminé par des signaux environnementaux comme la longueur du jour, l’abondance de nourriture… ou encore l’endroit où la larve tombe à la fin de son développement. C’est le cas chez le ver marin appelé la Bonellie verte, chez qui les larves naissent sans sexe déterminé, mais qui vont devenir femelles si elles tombent sur le sol, ou devenir mâles si les larves tombent… sur une femelle.

Précisons que la différence entre mâles et femelles est extrême, c’est à dire que si la larve devient femelle, elle atteindra sa forme adulte en 2 ans de développement, pour faire plus d’1m – alors que les mâles ne font que quelques millimètres, et leur seul objectif est de se coller à la trompe des femelles pour être aspirés jusqu’à leur appareil reproducteur (ce n’est pas sans rappeler comment marche la fécondation chez les plantes d’ailleurs)… ou chez les baudroies chez qui les mâles restent tout petits et fusionnent avec l’épiderme et les vaisseaux sanguins de la femelle pour se transformer en sorte de testicules parasitaires.
Bref, toutes ces modalités de déterminisme du sexe ont des avantages et des défauts et ont été sélectionnées spécifiquement pour répondre à des contraintes écologiques particulières, comme par exemple le niveau de difficulté de trouver un partenaire sexuel.
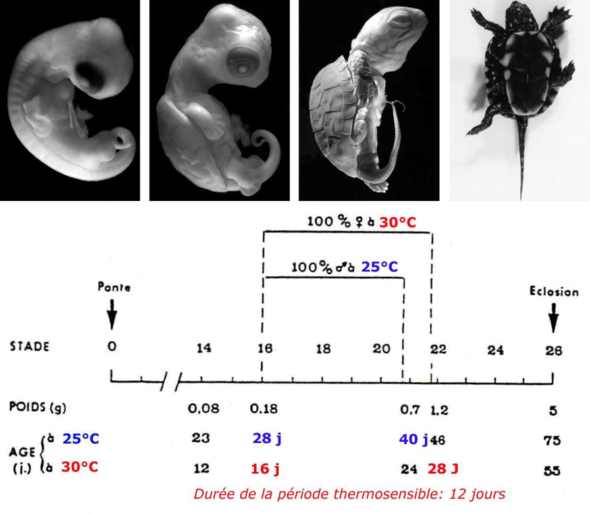
Puisque ce qui nous intéresse aujourd’hui dans cette émission, c’est ce que les gènes ne déterminent pas, je vous propose de nous concentrer ici sur la détermination du sexe par l’environnement, et notamment par la température et je vais commencer par évoquer ce qu’il se passe chez une espèce de tortue : la cistude d’Europe.

Chez cette espèce (protégée), si on incube les œufs à 27°C on obtient uniquement des mâles à l’éclosion. Par contre ce ne sont que des femelles qu’on obtient si l’on incube ces œufs à 30°C. En affinant ces résultats à l’aide d’expériences d’incubation à des températures très précises, on s’aperçoit qu’il existe une gamme de températures de transition qui fournit des mâles et des femelles. On appelle cette gamme de température la TRT pour Transitional Range of Temperature (l’intervalle de température de transition). Et il y a aussi une température où le sex ratio obtenu est de 50% et qu’on appelle la température pivot.
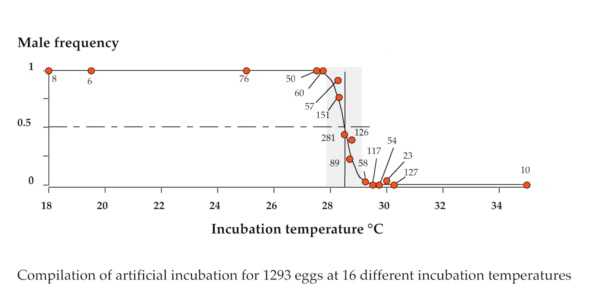
L’existence de la TRT et d’une température pivot a longtemps laissé les chercheurs perplexes (et à vrai dire continuent de le faire). Car quand on obtient un sex ratio de 50%, ça évoque tout de suite un déterminisme sexuel génétique et plus particulièrement qui exploite des chromosomes sexuels. On appelle ça un déterminisme hétérogamétique. Nous les humains, et quasi tous les mammifères, on a un déterminisme hétérogamétique de type XY où le chromosome Y détermine le sexe mâle. Chez d’autres animaux, comme par exemple les oiseaux, il existe un déterminisme hétérogamétique de type ZW où le chromosome W détermine le sexe femelle (donc les mâles sont ZZ). Le truc un peu bizarre, c’est qu’on trouve des chromosomes Z et W chez de nombreuses tortues, et X et Y chez d’autres, et pourtant plus de 100 espèces de tortues ont un déterminisme sexuel par la température. Donc, ce que la température pivot semble évoquer, c’est qu’à celle-ci, le système hétérogamétique de la cistude peut déterminer le sexe. Par contre, quand on s’écarte de la température pivot, c’est l’environnement qui décide…
Y’a aussi quelque chose à prendre en compte, c’est que par rapport aux tortues, les mammifères et les oiseaux sont des espèces endothermes, qui produisent leur propre chaleur (Un petit rappel de l’émission 315 sur l’écophysiologie). Les tortues, comme la plupart des sauropsidés (le nom plus exact pour dire reptile) sont des ectothermes et dépendent donc du milieu externe pour changer de température interne.
Et l’environnement décide le sexe uniquement à un moment précis de l’embryogenèse des cistudes.
Ça veut dire que vous aurez bon congeler ou cuire vos cistudes après l’éclosion, elles ne changeront pas de sexe (ne le faites pas svp, on rappelle que c’est une espèce protégée).
Ce moment, c’est celui du développement des gonades, et l’étude du développement de ces structures a révélé que presque tous les animaux partent d’une structure embryonnaire qui peut potentiellement donner soit des testicules, soit des ovaires, mais part d’une structure bi-potentielle indéterminée. En gros, on part avec une gonade asexuée qui, en fonction du type de déterminisme, va intégrer des signaux environnementaux ou génétiques pour diriger le développement de l’organe vers celui de testicules ou d’ovaires.
Ça on peut notamment s’en rendre compte en réalisant des expériences hardcore de culture de gonades embryonnaires au laboratoire, ce qui est faisable notamment chez les tortues de Floride, Trachemys scripta elegans.

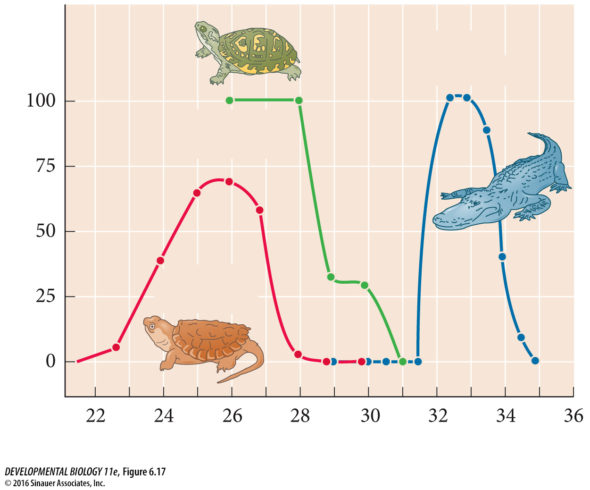
Chez cette espèce, lorsque les œufs d’une tortue sont incubés à 26°C, on obtient que des mâles et lorsque cette température monte à 32°C, on obtient que des femelles. C’est vraiment très proche du système de la Cistude quoi. Mais c’est cette tortue de Floride qui a été choisie par plusieurs laboratoires comme organisme modèle pour explorer la génétique de sa détermination sexuelle. Et le moins qu’on puisse dire, c’est que c’est pas fastoche! Chez une espèce chez qui il y a un déterminisme sexuel génétique, c’est assez pratique d’identifier les gènes qui sont responsables de cette détermination : vous identifiez les gènes qui se trouvent sur les chromosomes sexuels. Vous identifiez ceux qui présentent des expressions différentes chez un embryon mâle et un embryon femelle, puis vous les altérez et identifiez ceux qui ont un impact sur la formation des gonades. J’dis ça mais ça a dû prendre quelques dizaines d’années tout de même. Mais pour les espèces dont le déterminisme sexuel est lié à la température, tous les gènes sont potentiellement des cibles. Et c’est beaucoup plus sioux de détecter les variations d’expressions de gènes qui sont liés à la température ET qui ont un lien avec le déterminisme. Y’a trouze mille raisons pour lesquelles les gènes peuvent avoir des variations d’expression à la température et qui n’ont aucun lien avec le déterminisme sexuel. Pas étonnant que cela fait plus de 50 ans que la recherche faisait du sur-place sur l’étude de la détermination du sexe par la température.
Et bien il y a moins d’un an, des équipes sino-américaines ont fait un bond de géant dans la compréhension du déterminisme sexuel par la température chez la tortue de Floride. Et si le sujet est pertinent par rapport au thème de l’émission de ce soir, c’est qu’il existe un lien étroit et surprenant entre l’épigénétique, et la manière dont on obtient des filles et des garçons chez ces tortues.
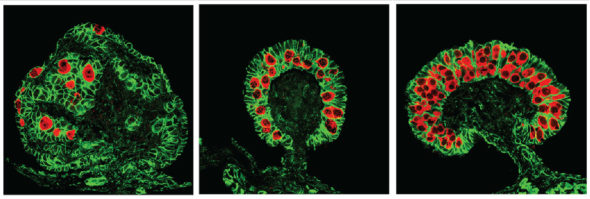
En fait, cela faisait des années que plusieurs équipes se disaient que s’intéresser aux différences d’expression des gènes ne suffirait pas à comprendre le phénomène de la détermination du sexe par la température. Ces équipes ont commencé par caractériser l’état des molécules d’ADN dans des gonades en formation, cultivées à différentes températures. Je laisserai à Eléa le soin d’expliquer cet état des molécules d’ADN, mais, en gros, ça revient à se demander si l’ADN est plus ou moins compacté et décoré à certains endroits (un peu comme un arbre de Noël avec des guirlandes et des boules quoi). Ça, c’est typiquement un sujet d’épigénétique. Mais encore une fois, trouver des différences de décoration et compaction de l’ADN dans des cellules de gonades cultivées à différentes températures, cela ne permet pas de dire que ces différences sont à l’origine du destin que va prendre ces gonades. Corrélation, n’est pas causation, vous connaissez tout le tintouin.
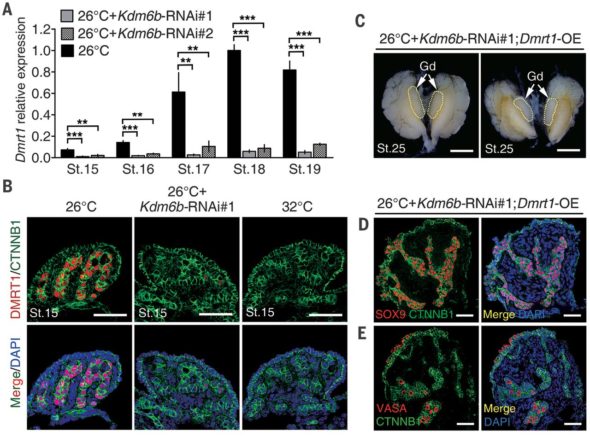
Là où ces équipes sino-américaines ont été futées, c’est qu’elles ont recoupé les informations entre l’état de l’ADN ET des différences d’expressions de gènes dans les gonades cultivées à différentes températures. Elles ont fait un pari entre des types de différences de décoration de l’ADN et les gènes qui sont responsables d’établir ces décorations. Parmi ces gènes, une enzyme portant le nom barbare de KDM6B et qui est une déméthylase de l’histone qui retire spécifiquement les trois groupements méthyles de la 27ème lysine de l’histone H3. WOW, merci le jargon. Plutôt que de se prendre la tête avec des noms barbares, jonglons plutôt avec des concepts. Ces équipes ont trouvé un gène qui produit une protéine décoratrice un peu maniaque, qui fait le ménage sur l’ADN : KDM6B (que je vais appeler la protéine Mary Poppins. Cette protéine est produite à 26°C dans les gonades juste avant leur différenciation, et n’est pas produite à 32°C. Qui plus est, Mary Poppins ne semble pas du tout apprécier les changements de températures. Si on commence à cultiver les gonades à 26°C, Mary Poppins commence à être produite, mais si on change brusquement la température d’incubation à 32°C, ça s’arrête.
La prouesse réalisée par ces chercheurs, c’est qu’ils ont trouvé un moyen d’empêcher la production de Mary Poppins (KDM6B) même quand on incube à 26°C. Je ne développerai pas cette technique (qui s’appelle l’interférence à ARN (et qui sera le sujet d’un prochain épisode !), mais vous pouvez me croire sur parole quand je vous dis que c’est chaud patate de développer ce genre de protocoles chez des espèces bizarres comme des tortues de Floride.
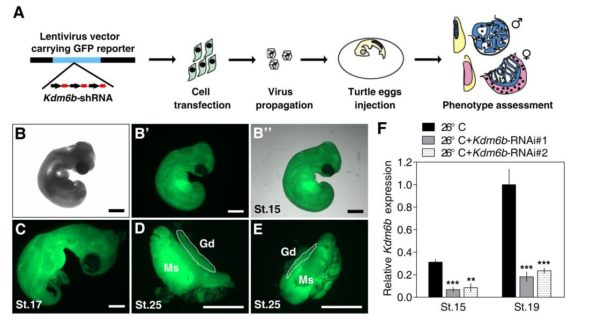
Et bien en empêchant Mary Poppins de ramener le bout de son parapluie magique dans des embryons incubés à 26°C (et qui devraient normalement donner des mâles), les chercheurs ont observés que les gonades obtenues ressemblaient à celles classiques de femelles. C’est l’une des premières preuves montrant qu’il faudrait avoir notre Mary Poppins dans les gonades de tortues pour obtenir des mâles. Mais ils sont allés plus loin! Ils ont réussi à voir où pouvait agir Mary Poppins (en gros où est-ce qu’elle changeait la décoration de l’ADN) et en déduire quels pouvaient être les gènes affectés par son action (ou inaction). Je vous la fais courte : ils ont identifié un gène qui s’appelle Dmrt1. Dans l’ADN de la tortue, l’endroit où on trouve Dmrt1 est bel et bien un endroit où Mary Poppins aime faire le ménage. Quand Mary Poppins fait le ménage sur le gène Dmrt1, celui-ci peut s’exprimer fortement et produire des protéines Dmrt1.
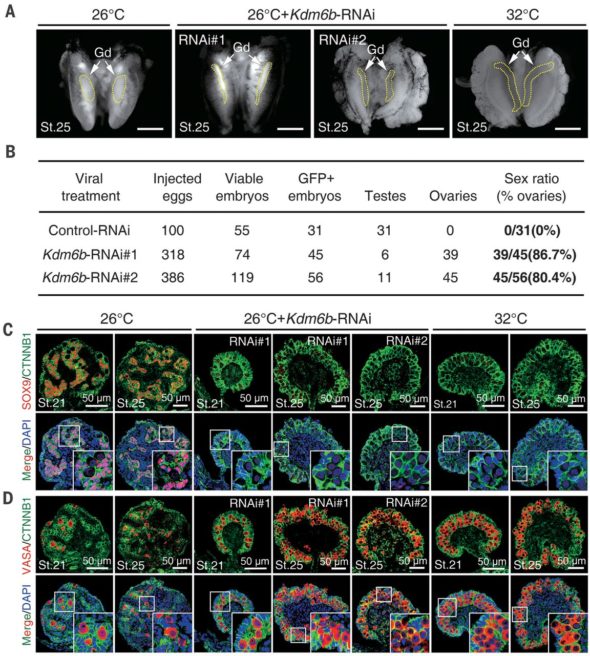
Mais en plus, ils ont réalisé une expérience dans laquelle ils empêchent à Mary Poppins de venir à 26°C, tout en rajoutant artificiellement des protéines Dmrt1. Et là, je vous le demande, à votre avis qu’est-ce que ça pourrait bien donner comme gonades finales?
Tout cela semble donc très fortement suggérer que ces équipes ont découvert des gènes du déterminisme sexuel par la température, et que le mécanisme derrière cet étrange phénomène, s’appuie sur de l’épigénétique. J’espère donc que cette première partie vous aura fortement titillé votre curiosité, avant de nous plonger maintenant dans les explications d’Eléa sur ce qu’est réellement l’épigénétique.
Eléa vous parle du monde merveilleux de l’épigénétique.
Dans chacun des noyaux de chacune de nos cellules, il y a près de 2m d’ADN à stocker. On parle ici donc de cellules humaines dont le patrimoine génétique représente près de 3 milliards de nucléotides (A, T, G et C) répartis sur 23 paires de chromosomes. 2m, dans un compartiment de 10µM environ. Vous vous imaginez ? Pour réaliser cette prouesse, la cellule dispose de tout un tas de protéines autour desquelles l’ADN s’enroule, de façon la plus compacte possible.

Du coup, on en a déjà parlé dans l’épisode 355 entre autres, mais l’ADN est un enchaînement de deux molécules, formées de petites briques moléculaires (les nucléotides), enroulés en double hélice, et qui forment en plus un enroulement compact, comme les câbles à ressorts des vieux téléphones… (un temps que les moins de 20 ans ne peuvent pas connaître). Sous cette forme : ADN+protéine bien agencés, on parle de chromosomes condensés. C’est une méthode de compaction extrêmement efficace, qui permet la séparation et la répartition de l’information génétique entre deux cellules filles, pendant la division cellulaire (et uniquement pendant la division cellulaire) mais autant c’est pratique pour séparer les deux bras des chromosomes de façon équitable… autant c’est très embêtant pour lire l’information contenue sur la molécule d’ADN.

Les protéines chargées de la lecture doivent accéder physiquement à la séquence. Imaginez une pelote de laine : si vous voulez regarder un bout du fil sur lequel on aurait écrit quelque chose à l’intérieur, il faut d’abord dérouler la pelote pour pouvoir y accéder.
Si vous vous souvenez bien des épisodes précédents à ce sujet (et notamment celui sur la biologie moléculaire 355), ou de vos cours de biologie – pour être exprimé, l’ADN doit être “transcrit” en une nouvelle molécule, par un complexe de protéines qu’on appelle l’ARN polymérase. Cette machinerie est envoyée au bon endroit grâce à des protéines de ciblage qu’on appelle les facteurs de transcription, qui vont reconnaître des petites signatures d’ADN spécifiques juste avant le gène, dans des régions qu’on appelle les promoteurs, s’y lier, et recruter la polymérase. Il existe des facteurs de transcription activateurs (qui vont déclencher la lecture) – et il existe des facteurs de transcription répresseurs (dont la présence empêche l’accès à la séquence).
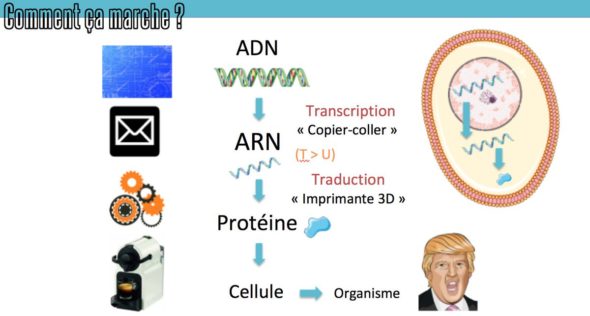
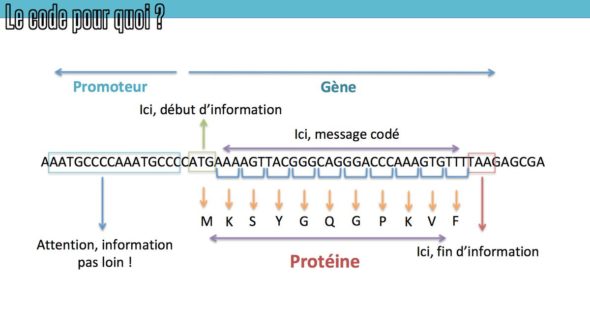
Pierre vous évoquait notamment le rôle important des facteurs de transcriptions dans l’épisode Evo-Devo 2, en soulignant l’importance en évolution des bouts d’ADN qui sont reconnus par ce genre de protéines. En effet, si des mutations font apparaitre ou disparaitre ces sites de fixation des facteurs de transcription, ça peut complètement changer l’expression des gènes, sans que les protéines changent elle-même.
Du coup, lorsque la cellule ne se divise pas, les chromosomes n’ont plus lieu d’être condensés, et on dit qu’ils se décompactent. Mais ce n’est pas pour autant que l’ADN redevient un long fil tranquille ! Non ! L’ADN reste enroulé autour de ces protéines à faire des pelotes, que l’on appelle des histones, en un complexe que l’on appelle le nucléosome. Un nucléosome, c’est une sorte de perle de 8 protéines histones : H2A, H2B, H3 et H4, autour desquelles l’ADN s’enroule d’exactement 146 paires de bases. Une cinquième histone, l’histone H1 sert en quelque sorte de “verrou” à la boucle d’ADN enfilée autour de sa perle d’histones. Ce mélange protéine/ADN bien organisé est ce qu’on appelle la chromatine.

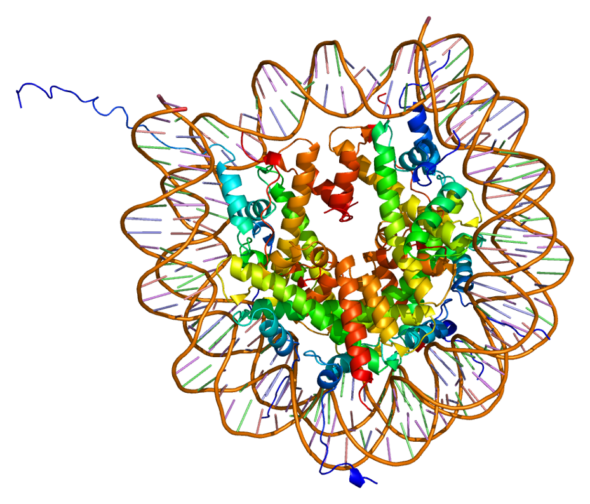
Autour de ces nucléosomes, l’ADN se répartit en territoires dans le noyau : des territoires plutôt compacts, dans lesquels on retrouve des gènes inaccessibles, très enroulés, inactifs, et des territoires plutôt moins compacts, déroulés, dans lesquels on retrouve des gènes accessibles, actifs qui sont exprimés. On parle souvent de chromatine “fermée”, ou de chromatine “ouverte”. On dit aussi “hétérochromatine” ou “euchromatine”.
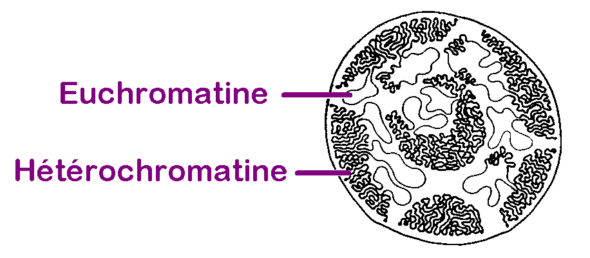

Pour continuer de filer la métaphore laineuse, c’est comme si on déroulait complètement une pelote, mais qu’ensuite on rangeait le tout dans une boite : on verrait qu’il y a des bouts de fils libres et des entrelacs spontanés de fils compactés.
Alors la frontière est floue entre ces territoires, et l’ensemble est assez dynamique. Et si on regarde plus près, ces territoires ont des caractéristiques particulières, et on retrouve des modifications chimiques différentes dans différentes régions du noyau de la cellule.
Dans la liste des modifications que l’on retrouve, on peut voir des modifications sur les histones (les protéines autour desquelles s’enroule l’ADN), ou des modifications directement sur les bases de l’ADN, dont ce qu’on appelle la méthylation de l’ADN (ajout d’un groupement méthyle : 1 carbone, 3 hydrogènes, sur la base “C”, la cytosine). Ces petites modifications vont modifier la façon dont l’ADN et les protéines interagissent entre elles, et bloquer ou favoriser la liaison des protéines de lectures à des endroits différents.

Ainsi, pour modifier l’expression d’un gène, vous pouvez changer la séquence de l’ADN (A, T, C, G) et cette modification sera irréversible si elle n’est pas réparée (vous vous souvenez de l’épisode 361). Ca, c’est de l’ordre de la génétique.
Ou vous pouvez rajouter des décorations sur votre ADN, ou les protéines nécessaires au bon enroulement de l’ADN pour modifier les possibilités d’accès aux facteurs qui doivent lire vos gènes. C’est l’épigénétique. Comme vous n’altérez pas la séquence, mais seulement la façon dont elle est exposée, ces modifications sont plutôt réversibles, et peuvent dépendre de facteurs internes, comme de facteurs externes (comme la température, dont parlait Pierre tout à l’heure.)
Grâce à ces mécanismes, vous augmentez considérablement les possibilités d’utiliser une seule et même information génétique.
Ca vend du rêve non ? Alors en pratique, ça donne quoi ?
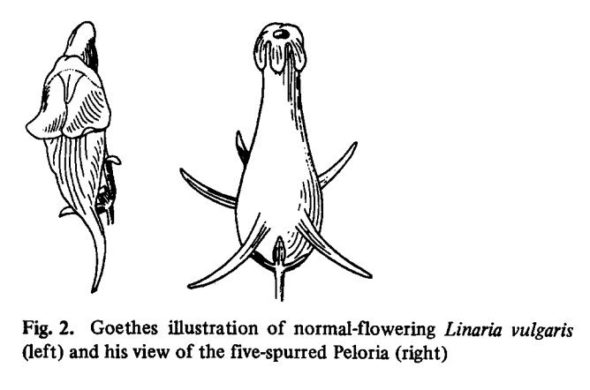
Je vous donne un exemple de conséquences de marques épigénétiques chez une plante. La linaire commune. Au 18ème siècle, un étudiant envoie à Linné une fleur monstrueuse, avec 5 pétales symétriques au lieu d’une forme de “gueule de loup”. Linné pense qu’il s’agit d’une espèce différente, mais des chercheurs montrent que non seulement il s’agit de la même espèce, mais que le gène responsable de la symétrie des fleurs (Lcyc) n’est pas muté. La séquence est normale, rien ne fait sens. Jusqu’à ce qu’ils se rendent compte qu’en fait, la partie précédent le gène, son promoteur est méthylé. Le gène ne peut ainsi plus être lu et la symétrie de la fleur change.
Vous pouvez lire cette histoire dans le détail dans un fil du plantoscope.

Epigénétique et détermination du sexe :
Comme vous le disait Pierre tout à l’heure à propos de la tortue : A 26°C, la cellule de tortue est capable de produire la protéine Mary Poppins (KDM6B vous vous souvenez). Cette protéine est capable d’enlever une petite modification, la méthylation, sur l’histone H3. Conséquence : ce petit changement rend les alentours du gène Dmrt1 (un gène crucial pour lancer le développement des organes mâle) un peu plus accessibles à des facteurs de transcription. Alors qu’il est “caché” d’ordinaire, il va être exprimé à ce moment là, et déclencher une réaction en chaîne de lecture de gènes, qui vont permettre de mettre en place le sexe masculin de notre petite tortue. Si c’est pas beau ?
Alors il y a d’autres mécanismes épigénétiques amusants qui ont lieu dans certaines espèces de mammifères. Par exemple, le fait que nous les filles avons deux chromosomes X, et que les garçons n’aient qu’un X de leur mère, et un Y de leur père pose un problème : comme les deux chromosomes X sont identiques, cela veut dire qu’on a deux fois la copie d’un même gène.
Si ce même gène est exprimé pareil depuis ses deux copies, on aurait donc systématiquement deux fois plus d’une même protéine que les garçons. C’est pas très égalitaire vous allez me dire… mais rassurez vous, nos cellules sont capables d’inactiver une des copies du X. Comment ? Hé bien le chromosome X porte un gène intéressant qui ne code pas pour une protéine, mais pour un ARN appelé Xist (X inhibitory specific transcript). Lorsqu’il est exprimé, il recouvre totalement l’exemplaire du X duquel il est exprimé, le rendant inactif. Pendant le développement, l’un des deux gènes Xist (et seulement l’un des deux, au hasard) est méthylé, le rendant inaccessible à la machine de lecture. Comme il n’est pas exprimé, le produit du gène ne recouvre pas son chromosome, et donc on obtient un chromosome X actif, et un X inactif. Bingo. Plus d’informations ici.
Chez le chat, sur le chromosome X, il existe un gène qui a deux variants, gouvernant la couleur orange, ou la couleur noire respectivement. Lors du développement embryonnaire, on part d’une seule cellule qui se divise en tout un tas d’autres cellules filles pour former l’ensemble des tissus. Pour les mâles, il n’y a qu’une version de ce gène de la couleur, et donc ils seront forcément entièrement roux, ou entièrement noirs. Par contre chez les femelles, comme on peut avoir deux versions du X, donc deux versions du gène de la couleur (orange ou noir), et que les copies du X sont inactivées aléatoirement dans chaque cellule, on peut se retrouver avec une chatte “écaille de tortue” ou “calico”, qui a des patches noirs, des patches oranges à la fois, répartis aléatoirement sur le corps.

Et ce mécanisme existe aussi sur d’autres chromosomes que le X. En réalité, les copies de gènes qui nous viennent du père ou de la mère ne sont pas exprimées pareil… C’est ce qui explique que si vous croisez un âne mâle avec une jument, vous obtenez une mule, alors que si vous croisez une ânesse avec un cheval, vous obtenez un bardot, qui ne ressemble pas du tout au mulet.

Ou que si vous croisez un lion avec une tigresse, vous obtenez un ligre, alors que si vous croisez une lionne avec un tigre, vous obtenez un tigron…

Dans tous les cas, la descendance à des aspects très différents. Pendant le développement embryonnaire, une seule version des gènes (paternels ou maternels). Et cela s’explique par le mécanisme de l’”empreinte parentale”. Lors de la formation des gamètes – donc soit les ovules, soit les spermatozoïdes, une mosaïque de marques épigénétique, dont principalement de la méthylation sur les cytosines, se met un peu partout… enfin, un peu partout mais sur des endroits précis. Certains gènes maternels ou paternels sont inactivés pour éviter un “surdosage” de gène, et les deux versions des gènes ne sont pas lues de la même manière pendant le développement.
Une des conséquences observables de ce phénomène, c’est que si l’on change l’imprinting (empreinte) de la version maternelle ou paternelle de la même région du chromosome 15, on se retrouve avec deux maladies différentes :
- Le syndrome de Prader Willi, qui provoque une obsession pour la nourriture, un retard mental, des difficultés d’apprentissage…
- Alors que le syndrome d’Angelman est plutôt caractérisé par des enfants avec un retard mental, des difficultés de motricité, l’absence de parole, mais on les reconnaît parce qu’ils sont souvent très gais et qu’ils sourient beaucoup.
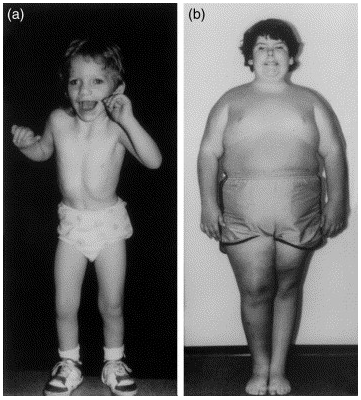
Ce phénomène d’empreinte concerne a priori 150 gènes chez la souris, 100 chez l’homme (on en a grosso-modo 20 000) et une douzaine chez les plantes à fleurs. Et ce mécanisme n’existe que chez les mammifères et les plantes !
Le pourquoi du comment ? Personne ne sait exactement… et c’est le sujet de nombreuses théories évolutives et de nombreuses recherches. A quel point les gènes soumis à empreinte sont-ils importants pour le développement, la survie d’une espèce ? Là on est à la frontière de ce qui est connu en biologie.
Mais la méthylation existe aussi sur des séquences d’ADN dont on sait qu’elles ont la capacité de se recopier et de “sauter” à de nouveaux endroits, les transposons. On connaît ces séquences depuis les années 50, grâce à Barbara McClintock qui travaillait sur la couleur du maïs. Ces séquences mettent un peu le bazar dans le génome. Oui, imaginez un bout d’ADN qui s’intègre au milieu d’un gène super important… comme une rature au milieu d’une phrase, vous ne pourrez plus lire la bonne information. Pour éviter ça, les transposons sont “mis en silence” grâce à la méthylation.

Epigénétique et différenciation cellulaire/développement :
Ces petites modifications ont donc de grosses conséquences sur le développement des organismes. Puisqu’une fois que le sexe de votre embryon est déterminé, et qu’on a choisi quels gènes de quels chromosomes s’expriment ou non, à quel moment, on lance une réaction en chaîne qui va permettre de … faire des organismes.
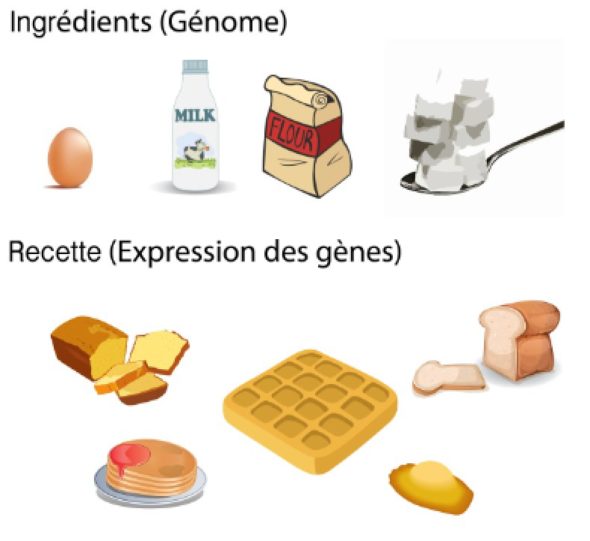
Pour vous faire une comparaison très marquante, imaginez vous que l’ensemble de vos gènes soient les mêmes ingrédients d’une recette de cuisine : des oeufs, du lait, du sucre, de la farine pour fabriquer un gâteau. L’épigénétique, c’est le fait de changer la recette et d’ajuster chacune des quantités des ingrédients principaux pour faire au choix un quatre quart, une crêpe, des madeleines, des gaufres…
Grâce à ces mécanismes, la cellule peut emprunter des chemins différents. C’est ainsi qu’à partir d’une cellule de départ qui a toutes les possibilités, la cellule souche, on peut générer toutes les populations de cellules ultérieures qui vont se différencier pour devenir chacun de nos organes. Il faut s’imaginer que notre première cellule est une bille en haut d’une montagne dont la pente est accidentée et vallonnée. De petits changements extérieurs (un peu de vent, un petit caillou, un petit objet qui entre en collision) peut lui faire emprunter une toute autre voie pour dévaler la pente de la montagne, jusqu’à arriver à son état final, le bas de la pente, duquel elle ne pourra plus jamais revenir.
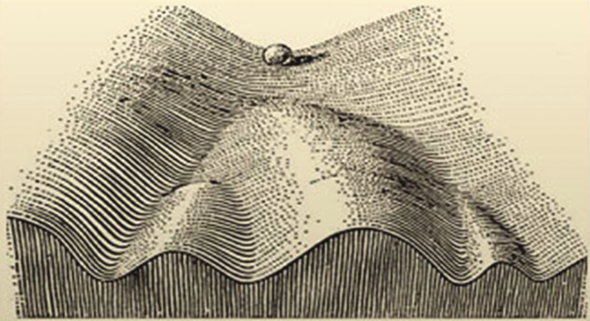
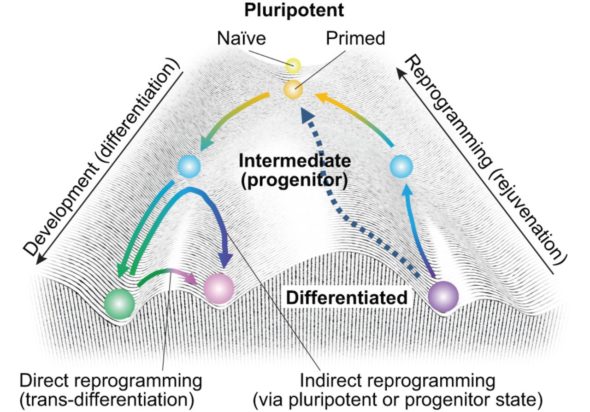
Cette image nous vient de Conrad Waddington, qui a proposé ce concept de “paysage épigénétique” et cette image que je viens de vous décrire. Une fois que l’on sait quelles marques épigénétiques et quels facteurs sont associés à tel type cellulaire…. Il n’est pas exclu que l’on puisse reprogrammer des cellules qui sont dans un état, pour les faire passer d’un autre état.

Les premières tentatives dans ce sens, ce sont les expériences de Yamanaka, qui a identifié quatre facteurs de transcription (donc les têtes de lecture de l’ADN). Normalement très exprimés dans les cellules embryonnaires, et plus du tout dans les cellules différenciées, il est possible de faire en sorte qu’une cellule “en bas de la pente” repasse dans un état de cellule souche en exprimant uniquement ces quatre facteurs ! Ces facteurs “maîtres” sont aussi appelés “facteurs pionniers” et peuvent aller chercher de l’information très inaccessible et programmer la séquence d’activation des gènes.
Et en parlant de reprogrammation, un autre exemple intéressant est celui de l’abeille. Les ouvrières sont petites, vivent entre 15 et 140 jours, ont des pattes adaptées à la collecte de pollen, des glandes spécialisées pour la fabrication de la cire… alors que les reines sont plus grosses, peuvent vivre jusqu’à 8 ans et surtout… sont fertiles. Elles ont des différences comportementales, cognitives… et pourtant la seule différence entre ces deux types d’abeille est leur nourriture à l’état larvaire. Les reines étant nourries à la gelée royale, alors que les futures ouvrières non. La gelée royale contient entre autres du phényl butyrate, qui est un inhibiteur connu d’enzymes HDAC: histone déacétylases, des protéines chargées de changer le “paysage épigénétique” et la compaction de la chromatine. Et si on analyse leurs différences épigénétiques, leurs profils de méthylation et la compaction de la chromatine à des endroits clés diffère effectivement. En plus, elle contient des protéines comme la Major Royal Jelly Protein 1 (MRJP1), aussi appelée Royalactine. Des études ont montré que cette protéine aidait à maintenir les cellules dans un état de pluripotence, c’est à dire qui gardent toutes leurs possibilités.
Alors attention, si la gelée royale rend l’abeille plus… royale, ça ne veut pas dire qu’en manger vous fera le même effet, allongera votre durée de vie grâce à un super effet épigénétique, contrairement à ce que vous dit le marketing des vendeurs de gelée royale. C’est super séduisant, mais… ça ne marche pas tout à fait comme ça.
Epigénétique et maladies :
Les modifications épigénétiques sont “visibles” dans la cellule depuis assez peu de temps, mais les technologies actuelles nous permettent d’avoir accès à des informations comme la position des histones, la fréquence leur modification, de méthylation à l’échelle de tout le génome. On peut maintenant comparer ces modifications dans différents organismes, dans des cellules différentes, dans des maladies, des tumeurs… et le domaine est en plein expansion. On dispose même de bases de données gigantesques qui compilent ces informations.
On sait désormais que les cellules cancéreuses ont des profils de méthylation complètement aberrants, que l’expression de certains facteurs de transcription (les gènes qui lisent l’ADN) peuvent être altérés dans certains types de cancer plus que d’autres. De nombreux projets de recherche essayent donc de comprendre quels sont les choses qui déconnent et comment une cellule “normale” devient cancéreuse, avec l’espoir de pouvoir inverser la tendance…
Attention aux dérives :
La question qui subsiste, c’est est ce que ces marques épigénétiques sont transmises à la descendance ? Est-ce que notre mode de vie et des facteurs extérieurs peuvent impacter nos générations futures ? Des marques comme la méthylation sont recopiées et maintenues à chaque division cellulaire, mais à part pour quelques gènes, le profil de méthylation est remis à zéro directement après la fécondation, et les marques sont remises en place pendant le développement.
Si on a très peu de recul sur ce qu’il peut se passer chez l’humain, car les générations sont longues, on sait en revanche que lorsqu’on stimule la perception d’une odeur chez une souris, le récepteur olfactif correspondant peut être surexprimé 2 générations plus tard. Ce qui laisse penser que certains effets puissent être transmis à la descendance.
Mais attention aux allégations qui disent que tous vos maux s’expliquent par les comportements ou le vécu des générations qui vous ont précédé.
Il existe plusieurs articles sur internet qui rapportent des associations entre une famine à telle ou telle période, et les statistiques de décès plusieurs générations plus tard.
La plus célèbre, souvent reprise par les médias comme “c’est prouvé, votre régime alimentaire influencera vos petits enfants” est l’une des études concernant le village d’Overkalix en Suède, pour lequel on dispose de 150 ans d’archives de récoltes et d’archives médicales. L’étude montre que les hommes dont le grand-père a vécu pendant une période de faibles récoltes entre 9 et 12 ans (on pré-suppose qu’il était donc mal nourri à ce moment là) ont un taux de mortalité deux fois plus faible que ceux qui ont vécu leurs 9 à 12 ans avec de bonnes récoltes, avec un taux plus élevé de maladies cardio-vasculaires et de diabète. Une corrélation que les auteurs expliquent par des mécanismes épigénétiques, CQFD. Ils n’apportent aucune preuve expérimentale qu’il existe une différence épigénétique (on a plus d’échantillons de ces gens à leurs 9 ans pour comparer avec leurs petits enfants, eux mêmes morts) et on ne prend pas en compte les changements d’habitudes alimentaires qui ont pu s’opérer en 150 ans et 2 générations… Donc s’il y a quelque chose, ce n’est certainement pas cette étude qui le montre. Pas mal de biais qui font que l’épigénétique n’explique pas tout non plus, même si on aimerait très fort.
Ce qu’il faut retenir, c’est que ces mécanismes épigénétiques permettent aux organismes de s’adapter très rapidement à quelque chose, dont l’environnement – (c’est grâce à des mécanismes épigénétiques que les plantes peuvent déclencher leur floraison, en intégrant des informations comme la température, l’humidité, la durée de la luminosité…) , de faire plus que ce qui est simplement codé physiquement, et que les changements qu’ils induisent, peuvent dans une certaine mesure, se transmettre à court terme. Tout n’est pas strictement génétique, et la régulation des gènes est un tantinet plus complexe que ce qu’on apprend à l’école.
Donc définitivement, gras, sexe, poils… les gènes ne suffisent plus pour expliquer tout ça.
Pour aller plus loin :
Un cours avec plein d’exemples de ce qu’est l’épigénétique.
Articles Scientifiques:
Ashman, T., Bachtrog, D., Blackmon, H., Goldberg, E., Hahn, M., Kirkpatrick, M., Kitano, J., Mank, J., Mayrose, I., Ming, R., Otto, S., Peichel, C., Pennell, M., Perrin, N., Ross, L., Valenzuela, N., and Vamosi, J. (2014). Tree of Sex: A database of sexual systems Scientific Data, 1 DOI: 10.1038/sdata.2014.15
Bachtrog, D., Mank, J. E., Peichel, C. L., Kirkpatrick, M., Otto, S. P., Ashman, T.-L., . . . The Tree of Sex, C. (2014). Sex Determination: Why So Many Ways of Doing It? PLOS Biology, 12(7), e1001899. doi: 10.1371/journal.pbio.1001899
Ge, C., Ye, J., Weber, C., Sun, W., Zhang, H., Zhou, Y., . . . Capel, B. (2018). The histone demethylase KDM6B regulates temperature-dependent sex determination in a turtle species. Science, 360(6389), 645-648. doi: 10.1126/science.aap8328
Georges, A., & Holleley, C. E. (2018). How does temperature determine sex? Science, 360(6389), 601-602. doi: 10.1126/science.aat5993
Li, X. Y., & Gui, J. F. (2018). An epigenetic regulatory switch controlling temperature-dependent sex determination in vertebrates. Sci China Life Sci, 61(8), 996-998. doi: 10.1007/s11427-018-9314-3
S. Alexandra Garcia-Moreno, Michael P. Plebanek, Blanche Capel, Epigenetic regulation of male fate commitment from an initially bipotential system, Molecular and Cellular Endocrinology, Volume 468, 2018, Pages 19-30, ISSN 0303-7207, https://doi.org/10.1016/j.mce.2018.01.009 (http://www.sciencedirect.com/science/article/pii/S0303720718300194 )
Weber, C., & Capel, B. (2018). Sex reversal. Current Biology, 28(21), R1234–R1236. doi:10.1016/j.cub.2018.09.043
Tzika, E., Dreker, T. & Imhof, A. Epigenetics and Metabolism in Health and Disease. Front Genet9, (2018).
Liens:
Epigénétique :
L’épigénétique, quand l’environnement a son mot à dire sur la génétique. Parlons Peu, Parlons Science.
Il n’y a pas que les gènes dans la vie ! L’épigénétique, avec P’tite Jane — Science étonnante #54
L’épigénétique : il n’y a pas que les gènes dans la vie ! Science Étonnante
Epigénétique. Podcast Science
[BioTCom] 07 : épigénétique et chatons
[BioTCom-RS] 02 : retours sur l’inactivation du chromosome X
Epigénétique et Obésité : Et si la prise de poids était programmée avant la naissance ?
X Chromosome: X Inactivation . Scitable Nature Education
Détermination Sexuelle par la température :
Vertebrate sexual systems. Pleiotropy.
https://en.wikipedia.org/w/index.php?title=Temperature-dependent_sex_determination
There’s a genetic explanation for why warmer nests turn turtles female. Science News
How turning down the heat makes a baby turtle male. Phys.org
Les tortues marines se féminisent avec la hausse des températures. Est-ce grave ? The Conversation
Température, stress et sexe : le changement climatique va-t-il masculiniser les poissons ? The Conversation
How is the gender of some reptiles determined by temperature? Scientific American